Epoxide hydrolase 2
| View/Edit Human | View/Edit Mouse |
Soluble epoxide hydrolase (sEH) is a bifunctional enzyme that in humans is encoded by the EPHX2 gene.[3][4][5] sEH is a member of the epoxide hydrolase family. This enzyme, found in both the cytosol and peroxisomes, binds to specific epoxides and converts them to the corresponding diols. A different region of this protein also has lipid-phosphate phosphatase activity. Mutations in the EPHX2 gene have been associated with familial hypercholesterolemia.[3]
Tissue distribution
While most highly expressed in the liver, sEH is also expressed in other tissues including vascular endothelium, leukocytes, red blood cells, smooth muscle cells, adipocytes and the kidney proximal tubule.[4]
Catalyzed reactions
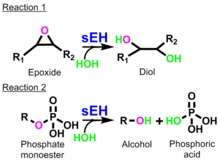
The form of sEH in the intracellular environment is a homodimer with two distinct activities in two separate structural domains of each monomer: the C-terminal epoxide hydrolase activity (soluble epoxide hydrolase: EC 3.3.2.10) and the N-terminal phosphatase activity (lipid-phosphate phosphatase: EC 3.1.3.76).[4] sEH converts epoxides, or three membered cyclic ethers, to their corresponding diols through the addition of a molecule of water.[4] The resulting diols are more water-soluble than the parent epoxides, and so are more readily excreted by the organism.[4]
The C-term-EH catalyzes the addition of water to an epoxide to yield a vicinal diol (reaction 1).[4] The Nterm-phos hydrolyzes phosphate monoesters, such as lipid phosphates, to yield alcohols and phosphoric acid (reaction 2).[4] The C-term-EH hydrolyzes one important class of lipid signaling molecules that includes many epoxyeicosatrienoic acids (EETs) that have vasoactive, anti-inflammatory and analgesic properties.[6]
sEH also appears to be the hepoxilin hydrolase that is responsible for inactivating the epoxyalcohol metabolites of arachidonic acid, hepoxilin A3 and hepoxiin B3.[7][8]
Discovery
The sEH was first identified in the cytosolic fraction of mouse liver through its activity on epoxide containing substrates such as juvenile hormone and lipid epoxides such as epoxystearate.[9] The soluble EH activity was shown to be distinct from that of the microsomal epoxide hydrolase (mEH) previously discovered with a different substrate selectivity and cellular localization than the mEH. Studies using a lipid epoxide as a substrate detected this activity in the soluble fraction of multiple organs, though at a lesser amount than in liver and kidney.[10] The enzyme activity was detected in rabbits, mice and rats, and humans, and it is now believed to be ubiquitous in vertebrates.[11] The proposed enzyme was first named cytosolic epoxide hydrolase; however, after its discovery inside the peroxisomes of some organs, it was renamed soluble epoxide hydrolase or sEH.[11]
Function
sEH has a restricted substrate selectivity, and has not been shown to hydrolyze any toxic or mutagenic xenobiotics.[4] Conversely, the sEH plays a major role in the in vivo metabolism of endogenous lipid epoxides, such as the EETs and squalene oxide, a key intermediate in the synthesis of cholesterol.[4] EETs are lipid signaling molecules that function in an autocrine and paracrine manner.[12] They are produced when arachidonic acid is metabolized by cytochrome p450s (CYPs).[12] These enzymes epoxidize the double bonds in arachidonic acid to form four regioisomers.[4] Arachidonic acid is also the precursor of the prostaglandins and the leukotrienes, which are produced by cyclooxygenases and lipoxygenases, respectively.[6] These lipids play a role in asthma, pain, and inflammation and are the targets of several pharmaceuticals.[13] The EET receptor or receptors have not been identified, but several tools for the study of EET biology have been developed, these include small molecule sEH inhibitors, EET mimics and sEH genetic models. Through the use of these tools, as well as the EETs themselves, the EETs have been found to have anti-inflammatory and vasoactive properties.[4] Several disease models have been used, including Ang-II induced hypertension and surgical models of brain and heart ischemia. In vitro models such as isolated coronary rings and platelet aggregation assays have also been employed.[4]
The proposed role of sEH in the regulation of hypertension can be used as a simple model of sEH function in the kidney.[14] Here the EETs are vasodilatory, and can be thought of as balancing other vasoconstrictive signals. sEH hydrolyzes the EETs to form the dihydroxyeicosatrienoic acids (DHETs).[14] These molecules are more water-soluble and are more easily metabolized by other enzymes, so the vasodilatory signal is removed from the site of action through excretion, tipping the balance of vasoconstrictive and vasodilatory signals towards vasoconstriction. This change in the lipid signaling increases vascular resistance to blood flow and blood pressure.[4] By reducing sEH epoxide hydrolase activity, and thereby shutting off the major route of metabolism of the EETs, the levels of these molecules can be stabilized or increased, increasing blood flow and reducing hypertension.[14] This reduction in sEH activity can be achieved in genetic models in which sEH has been knocked out, or through the use of small molecule sEH inhibitors.[15]
This simplified model is complicated by a number of factors in vivo. The EETs display different properties in different vascular beds.[12] The DHETs are more readily excreted, but they have yet to be fully characterized, and may possess biological properties themselves, complicating the balance of signals described in the simplified model.[4] There are epoxides of other lipids besides arachidonic acid such as the omega three docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) epoxides.[16] These lipid epoxides have been shown to have biological effects in vitro in which they inhibit platelet aggregation.[17] In fact, in some assays they are more potent than the EETs.[18] Other epoxidized lipids include the 18-carbon leukotoxin and isoleukotoxin.[19] The diepoxide of linoleic acid can form tetrahydrofuran diols,[20]
sEH metabolizes the biologically active epoxyalcohol metabolites of arachidnoic acid, hepoxilin A3 (8-hydroxy-11S,12Sepoxy-(5Z,8Z,14Z)-eicosatrienoic acid) to trioxilin A3 (8,11,12-trihydroxy-(5Z,9E,14Z)-eicosatrienoic acid) and hepoxilin B3 (10-hydroxy-11S,12Sepoxy-(5Z,9E,14Z)-eicosatrienoic acid) to trioxlin B3 (10,11,12-trihydroxy-(5Z,9E,14Z)-eicosatrienoic acid.[21] These trihydroxy products are generally considered to be inactive and the sEH pathway is generally considered to limit the actions of the hepoxilins.[8][21]
The phosphatase activity of sEH has been shown to hydrolyze in vitro lipid phosphates such as terpene pyrophosphates or lysophosphatidic acids.[4] However, its biological role is still unknown.
Clinical significance
Through metabolism of EETs and other lipid mediators, sEH plays a role in several diseases, including hypertension, cardiac hypertrophy, arteriosclerosis, brain and heart ischemia/reperfusion injury, cancer and pain.[12] Because of its possible role in cardiovascular and other diseases, sEH is being pursued as a pharmacological target, and potent small molecule inhibitors are available.[15]
Because of the implications to human health, sEH has been pursued as a pharmaceutical target and several sEH inhibitors have been developed in the private and public sectors.[15] One such inhibitor, UC1153 (AR9281), was taken to a phase IIA clinal trial for treatment of hypertension by Arête Therapeutics.[22] However, UC1153 failed the clinical trial, due in large part because of its poor pharmacokinetic properties.[15] Since this trial, a different sEH inhibitor, GSK2256294, developed for chronic obstructive pulmonary disease by GlaxoSmithKline has entered the pre-recruiting phase of a phase I clinical trial for obese male smokers.[23] Thus, interest continues in sEH as a therapeutic target.
One indication of the possible therapeutic value of sEH inhibition comes from studies examining physiologically relevant single nucleotide polymorphisms (SNPs) of sEH in human populations.[24] The Coronary Artery Risk Development in Young Adults (CARDIA) and the Atherosclerosis Risk in Communities (ARIC) studies both associated SNPs in the sEH coding region with coronary heart disease.[25][26] In these studies, two nonsynonymous SNPs were identified, R287Q and K55R. R287Q changes the arginine in position 287 in the most frequent allele to glutamine, while K55R changes the lysine in position 55 to an arginine. R287Q was associated with coronary artery calcification in African American population participating in the CARDIA study.[25][27] The K55R allele is associated with the risk of developing coronary heart disease in Caucasians participating in the ARIC study, where it was also associated with a higher risk of hypertension and ischemic stroke in male homozygotes.[26]
References
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- 1 2 "Entrez Gene: Epoxide hydrolase 2, cytoplasmic".
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Morisseau C, Hammock BD (2013). "Impact of soluble epoxide hydrolase and epoxyeicosanoids on human health". Annu. Rev. Pharmacol. Toxicol. 53: 37–58. doi:10.1146/annurev-pharmtox-011112-140244. PMC 3578707
 . PMID 23020295.
. PMID 23020295. - ↑ Harris TR, Hammock BD (September 2013). "Soluble epoxide hydrolase: gene structure, expression and deletion". Gene. 526 (2): 61–74. doi:10.1016/j.gene.2013.05.008. PMC 3733540. PMID 23701967.
- 1 2 Spector AA, Norris AW (March 2007). "Action of epoxyeicosatrienoic acids on cellular function". Am. J. Physiol., Cell Physiol. 292 (3): C996–1012. doi:10.1152/ajpcell.00402.2006. PMID 16987999.
- ↑ Cronin, A; Decker, M; Arand, M (2011). "Mammalian soluble epoxide hydrolase is identical to liver hepoxilin hydrolase". The Journal of Lipid Research. 52 (4): 712–9. doi:10.1194/jlr.M009639. PMC 3284163. PMID 21217101.
- 1 2 Muñoz-Garcia, A; Thomas, C. P.; Keeney, D. S.; Zheng, Y; Brash, A. R. (2014). "The importance of the lipoxygenase-hepoxilin pathway in the mammalian epidermal barrier". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1841 (3): 401–8. doi:10.1016/j.bbalip.2013.08.020. PMC 4116325. PMID 24021977.
- ↑ Morisseau C, Hammock BD (June 2008). "Gerry Brooks and epoxide hydrolases: four decades to a pharmaceutical". Pest Manag. Sci. 64 (6): 594–609. doi:10.1002/ps.1583. PMID 18383502.
- ↑ Gill SS, Hammock BD (February 1980). "Distribution and properties of a mammalian soluble epoxide hydrase". Biochem. Pharmacol. 29 (3): 389–95. doi:10.1016/0006-2952(80)90518-3. PMID 7362652.
- 1 2 Newman JW, Morisseau C, Hammock BD (January 2005). "Epoxide hydrolases: their roles and interactions with lipid metabolism". Prog. Lipid Res. 44 (1): 1–51. doi:10.1016/j.plipres.2004.10.001. PMID 15748653.
- 1 2 3 4 Imig JD, Hammock BD (October 2009). "Soluble epoxide hydrolase as a therapeutic target for cardiovascular diseases". Nat Rev Drug Discov. 8 (10): 794–805. doi:10.1038/nrd2875. PMC 3021468. PMID 19794443.
- ↑ Smith WL, Urade Y, Jakobsson PJ (October 2011). "Enzymes of the cyclooxygenase pathways of prostanoid biosynthesis". Chem. Rev. 111 (10): 5821–65. doi:10.1021/cr2002992. PMC 3285496. PMID 21942677.
- 1 2 3 Imig JD (September 2005). "Epoxide hydrolase and epoxygenase metabolites as therapeutic targets for renal diseases". Am. J. Physiol. Renal Physiol. 289 (3): F496–503. doi:10.1152/ajprenal.00350.2004. PMID 16093425.
- 1 2 3 4 Shen HC, Hammock BD (March 2012). "Discovery of inhibitors of soluble epoxide hydrolase: a target with multiple potential therapeutic indications". J. Med. Chem. 55 (5): 1789–808. doi:10.1021/jm201468j. PMC 3420824. PMID 22168898.
- ↑ Wagner K, Inceoglu B, Hammock BD (November 2011). "Soluble epoxide hydrolase inhibition, epoxygenated fatty acids and nociception". Prostaglandins Other Lipid Mediat. 96 (1–4): 76–83. doi:10.1016/j.prostaglandins.2011.08.001. PMC 3215909. PMID 21854866.
- ↑ Arnold C, Konkel A, Fischer R, Schunck WH (2010). "Cytochrome P450-dependent metabolism of omega-6 and omega-3 long-chain polyunsaturated fatty acids". Pharmacol Rep. 62 (3): 536–47. doi:10.1016/s1734-1140(10)70311-x. PMID 20631419.
- ↑ Spector AA (April 2009). "Arachidonic acid cytochrome P450 epoxygenase pathway". J. Lipid Res. 50 Suppl (Suppl): S52–6. doi:10.1194/jlr.R800038-JLR200. PMC 2674692. PMID 18952572.
- ↑ Zheng J, Plopper CG, Lakritz J, Storms DH, Hammock BD (October 2001). "Leukotoxin-diol: a putative toxic mediator involved in acute respiratory distress syndrome". Am. J. Respir. Cell Mol. Biol. 25 (4): 434–8. doi:10.1165/ajrcmb.25.4.4104. PMID 11694448.
- ↑ Moghaddam M, Motoba K, Borhan B, Pinot F, Hammock BD (August 1996). "Novel metabolic pathways for linoleic and arachidonic acid metabolism". Biochim. Biophys. Acta. 1290 (3): 327–39. doi:10.1016/0304-4165(96)00037-2. PMID 8765137.
- 1 2 Pace-Asciak, C. R. (2015). "Pathophysiology of the hepoxilins". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1851 (4): 383–96. doi:10.1016/j.bbalip.2014.09.007. PMID 25240838.
- ↑ "NCT00847899". Evaluation of Soluble Epoxide Hydrolase (s-EH) Inhibitor in Patients With Mild to Moderate Hypertension and Impaired Glucose Tolerance. ClinicalTrials.gov. Retrieved 2013-05-04.
- ↑ "NCT01762774". A Study to Assess the Safety, Tolerability, Pharmacokinetics and Pharmacodynamics of Single Doses of GSK2256294 in Healthy Volunteers, and Single and Repeat Doses of GSK2256294 in Adult Male Moderately Obese Smokers. ClinicalTrials.gov. Retrieved 2013-05-04.
- ↑ Fornage M, Hinojos CA, Nurowska BW, Boerwinkle E, Hammock BD, Morisseau CH, Doris PA (October 2002). "Polymorphism in soluble epoxide hydrolase and blood pressure in spontaneously hypertensive rats". Hypertension. 40 (4): 485–90. doi:10.1161/01.HYP.0000032278.75806.68. PMID 12364351.
- 1 2 Fornage M, Boerwinkle E, Doris PA, Jacobs D, Liu K, Wong ND (January 2004). "Polymorphism of the soluble epoxide hydrolase is associated with coronary artery calcification in African-American subjects: The Coronary Artery Risk Development in Young Adults (CARDIA) study". Circulation. 109 (3): 335–9. doi:10.1161/01.CIR.0000109487.46725.02. PMID 14732757.
- 1 2 Lee CR, North KE, Bray MS, Fornage M, Seubert JM, Newman JW, Hammock BD, Couper DJ, Heiss G, Zeldin DC (May 2006). "Genetic variation in soluble epoxide hydrolase (EPHX2) and risk of coronary heart disease: The Atherosclerosis Risk in Communities (ARIC) study". Hum. Mol. Genet. 15 (10): 1640–9. doi:10.1093/hmg/ddl085. PMC 2040335. PMID 16595607.
- ↑ Wei Q, Doris PA, Pollizotto MV, Boerwinkle E, Jacobs DR, Siscovick DS, Fornage M (January 2007). "Sequence variation in the soluble epoxide hydrolase gene and subclinical coronary atherosclerosis: interaction with cigarette smoking". Atherosclerosis. 190 (1): 26–34. doi:10.1016/j.atherosclerosis.2006.02.021. PMID 16545818.
Further reading
- Sandberg M, Hassett C, Adman ET, Meijer J, Omiecinski CJ (September 2000). "Identification and functional characterization of human soluble epoxide hydrolase genetic polymorphisms". J. Biol. Chem. 275 (37): 28873–81. doi:10.1074/jbc.M001153200. PMID 10862610.
- Farin FM, Janssen P, Quigley S, Abbott D, Hassett C, Smith-Weller T, Franklin GM, Swanson PD, Longstreth WT, Omiecinski CJ, Checkoway H (November 2001). "Genetic polymorphisms of microsomal and soluble epoxide hydrolase and the risk of Parkinson's disease". Pharmacogenetics. 11 (8): 703–8. doi:10.1097/00008571-200111000-00009. PMID 11692079.
- Cronin A, Mowbray S, Dürk H, Homburg S, Fleming I, Fisslthaler B, Oesch F, Arand M (February 2003). "The N-terminal domain of mammalian soluble epoxide hydrolase is a phosphatase". Proc. Natl. Acad. Sci. U.S.A. 100 (4): 1552–7. doi:10.1073/pnas.0437829100. PMC 149870. PMID 12574508.
- Petruzzelli S, Franchi M, Gronchi L, Janni A, Oesch F, Pacifici GM, Giuntini C (March 1992). "Cigarette smoke inhibits cytosolic but not microsomal epoxide hydrolase of human lung". Hum Exp Toxicol. 11 (2): 99–103. doi:10.1177/096032719201100207. PMID 1349227.
- Papadopoulos D, Gröndal S, Rydström J, DePierre JW (May 1992). "Levels of cytochrome P-450, steroidogenesis and microsomal and cytosolic epoxide hydrolases in normal human adrenal tissue and corresponding tumors". Cancer Biochem. Biophys. 12 (4): 283–91. PMID 1423213.
- Yoshimura K, Hanaoka T, Ohnami S, Ohnami S, Kohno T, Liu Y, Yoshida T, Sakamoto H, Tsugane S (2003). "Allele frequencies of single nucleotide polymorphisms (SNPs) in 40 candidate genes for gene-environment studies on cancer: data from population-based Japanese random samples". J. Hum. Genet. 48 (12): 654–8. doi:10.1007/s10038-003-0096-1. PMID 14634838.
- Sato K, Emi M, Ezura Y, Fujita Y, Takada D, Ishigami T, Umemura S, Xin Y, Wu LL, Larrinaga-Shum S, Stephenson SH, Hunt SC, Hopkins PN (2004). "Soluble epoxide hydrolase variant (Glu287Arg) modifies plasma total cholesterol and triglyceride phenotype in familial hypercholesterolemia: intrafamilial association study in an eight-generation hyperlipidemic kindred". J. Hum. Genet. 49 (1): 29–34. doi:10.1007/s10038-003-0103-6. PMID 14673705.
- Gomez GA, Morisseau C, Hammock BD, Christianson DW (April 2004). "Structure of human epoxide hydrolase reveals mechanistic inferences on bifunctional catalysis in epoxide and phosphate ester hydrolysis". Biochemistry. 43 (16): 4716–23. doi:10.1021/bi036189j. PMID 15096040.